Return to Essays top page
Contact:
Put at its simplest, genetic inheritance means that the cells which make up any organism's body contain complex chemicals called DNA and RNA, which act as coded instructions to the cell to produce other, different chemicals which control how cells behave, and so how the organism develops. This genetic code is passed on from parent to offspring during reproduction.
Some primitive species reproduce by cloning, that is, by a single parent producing offspring which are copies of itself and which therefore have a genetic code identical to the parent's. In this case, the genetic material changes very slowly by "mutation", that is, by alterations in the genetic code caused by transcription errors or damage. In organisms which use sexual reproduction the offspring derive their genes from two parents, with roughly half their genetic material coming from each parent: this leads to a constant re-combination of different genes producing slightly different organisms each time. This means that the species can evolve much faster, because for any change in the environment there will be some individuals who are better at coping with it than the rest.
This glossary is set out in logical rather than alphabetical order: read straight through from start to finish it should provide a crash-course in basic genetics. The first section concentrates on the physical components of genetic inheritance; the second on the interactions of genes with each other and with the organism's observed characteristics; the third covers the way genes are re-assorted during reproduction and some of the ways this can go wrong, linkages between genes, and the various forms of twinning; and the fourth looks at some terms you may encounter in discussions of evolutionary genetics.
The molecule is normally found as a double helix – two strands coiled round each other, with their bases interlocking to zip the strands together. If there's a C on one strand, there has to be a G in the matching position on the other strand, and so on.
Other variants of RNA are used to e.g. transport amino acids to their correct position on the mRNA template.
The exception is the sex chromosomes, which are the main factor in determining gender: in mammals females inherit two copies of the X chromosome, one from each parent, but males inherit an X chromosome from the mother and a Y chromosome (different size and shape) from the father.
Note that there are plenty of things that can go wrong with the sex chromosomes, resulting in e.g. XXY males who are male and yet have two XXs. Since all mammalian embryos start off female-shaped and only develop male characteristics quite late on, it is also possible for e.g. hormonal problems in the womb to produce an individual who is female in hormones and external anatomy, and yet has male genes and male internal structures.
In both females and XXY males only one X chromosome per cell is active, the other being switched off. The switched-off X chromosome stains differently from the active one in microscope slide preparations, and is known as a Barr Body.
If the X-chromosomes from each parent carry different colour-genes, for certain colour-combinations this can result in a patchwork-coloured ("tortoiseshell") animal, with e.g. some cells expressing an active X with a red gene, giving rise to red patches, and some with an active X with a black gene producing black patches.
Once the spare Xs in a developing embryo switch off (and Barr bodies form), each cell gives rise to a spreading patch of daughter-cells each of which has the same X switched off as in the original. The exact stage of embryological development at which this happens varies. If Barr bodies form very early in embryological development, when there are only a few cells in the embryo, each of those early cells then gives rise to quite a large area of the animal's body all with the same Xs switched on and off - hence if it is tortoiseshell it will be covered in big solid patches of colour. If Barr bodies form a bit later, when there are thousands of cells, then each will result on only a small patch of cells with that particular on/off combination, and the animal, if tortoiseshell, will be covered in subtle flecks of colour rather than patches.
The non-sexual chromosomes are referred to as the "autosomal" chromasomes.
Sperm cells do not contain mitochondria (they are too small) but egg-cells do: therefore, mitochondria are always inherited from one's mother. [In the case of cloned cells, where new genetic material has been inserted into an emptied ovum, they will be inherited from the female who donated the ovum, rather than the donor of the main genetic material.]
In addition to their own mitochondria, plants also have similar symbiotic bacteria called chloroplasts which likewise have their own separate DNA, and which carry out the photosynthesis which feeds the plant.
The exception again is the sex chromosomes: the Y chromosome is smaller than the X and also has some loci unique to itself, so many of the loci on the X chromosome do not have a matching position on the Y, and there are a few on the Y which don't have a match on the X. Hence male mammals, who pair an X with a Y, have many unpaired loci and are more likely to express recessive genes on the X chromosome than females are.
The mechanism of dominance is usually that the dominant allele produces a lot of a particular protein, while the recessive allele makes a weaker version of that protein, or less, or none. If we say that allele X codes for protein Y which has an effect Z on the phenotype of the organism, it often happens that allele X produces so much of protein Y that even a single copy of X is enough to trigger the effect Z, and so an organism with two X alleles and one which has one X allele and one modified allele x will develop just the same, even if the x allele produces little or no protein Y. Hence, it will not be possible to tell just by looking at the organism whether it has XX at that locus, or Xx, and X is said to be dominant and x recessive. Only if the organism has a double recessive xx pair, producing little or none of protein Y, will the phenotype be affected.
Where there are several possible alleles for a given locus, there is likely to be a hierarchy of dominance. There will be one gene which is dominant to all the rest, one which is recessive to all the rest and genes in the middle which are dominant to some of the others in the sequence and recessive to others. So for example ch is recessive towards C but dominant to ca (albinism).
This produces two daughter cells which each contain two of each type of chromosome (except for the XY pair in males), and have a genetic make-up identical to that of each other and of the original cell.
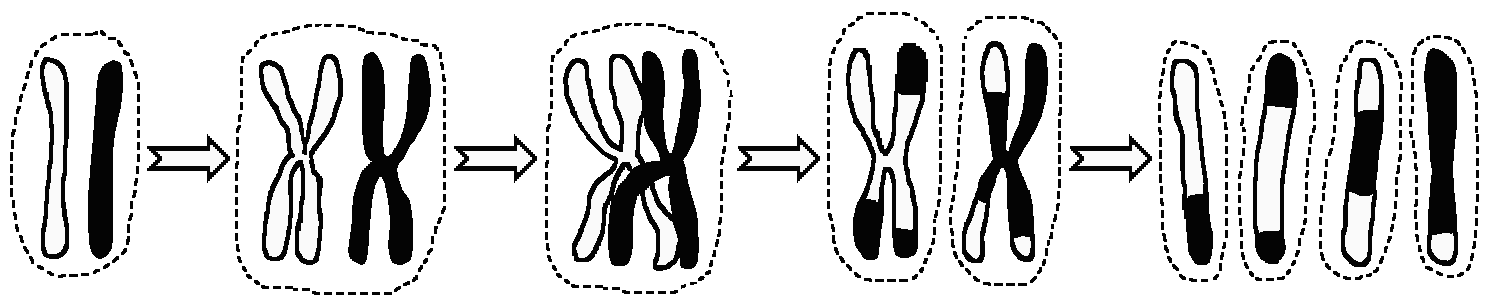
The chromosomes then separate again, breaking their chromatids apart at the cross-links in such a way that pieces of chromatid get mix-'n'-matched. You start with two chromosomes and four chromatids and you finish with two chromosomes and four chromatids, but the final chromatids often contain a mixture of material from both of the original chromosomes, and therefore from both of the organism's parents.
The cell then partially divides into two joined cells each of which normally contains only one of each type of paired chromosome, plus either an X or a Y sex chromosome. These cells are haploid (because they contain only one of each type of chromosome) but each of their chromosomes is still in the form of a pair of fused chromatids. The paired chromatids then part to form pairs of new chromosomes, and the cell then divides and separates these chromosomes into haploid cells again.
As a result one original diploid cell produces four haploid daughter cells, each containing one of each type of chromosome, with many or most of those chromosomes consisting of a mix of material from both of the chromosomes of that type which were in the original, diploid cell.
A somatic mutation is one affecting non-sexual cells within an organism. This can lead to e.g. cancers, but has no effect on (is not passed on to) the organism's offspring, except possibly in rare cases by RNA-mediated inheritance.
When mutation occurs in sex-cells it is passed on to any offspring which may form from those cells, resulting in an embryo containing genetic elements not found in either parent. Animals with major mutations often die in utero, and those which do live may be sickly and/or malformed (plants are more tolerant of distortion). But sometimes a mutant organism will have characteristics which are neutral or even useful, and which may then spread through the population and become an established, normal variation.
Also, in the case of minor mutations there are chemical buffers involved in embryological development which can often steer the organism towards a normal phenotype, even if it has some odd genes. These buffers tend to fail if the developing embryo is subjected to serious environmental stress, with the result that minor mutations which have been carried in a population for many generations may only start to affect phenotype when that population is exposed to environmental changes which make it a good idea to try out some new variants to see if they fit the altered situation.
Mutation of individual genes occurs when the DNA sequence of a gene is damaged/changed, sometimes because the organism is old and its cell-repair mechanisms are failing, and sometimes due to exposure to solar or other radiation, viral infection or to a "mutagenic" chemical.
Some loci are fracture-points at which mutation is especially likely to occur. For example in humans achondroplasia (a type of dwarfism) is dominant and yet normally occurs in children of normal-sized parents, the achondroplasia gene having occurred/recurred afresh in the sex-cells of one of the parents.
Chromosomal abnormalities normally occur when cell reproduction goes a bit wrong. During meiosis, chromatids may fail to separate symmetrically at the cross-linking stage, so that one chromatid ends up with a piece missing and one with a duplicate section: Down's Syndrome is due to such a duplication.
Alternatively the distribution of chromosomes between the daughter cells can go wrong, resulting in one cell with a whole chromosome missing and one with a duplicate chromosome. This happens during either mitosis or meiosis when chromosome pairs assort wrongly and both end up on the same side of the dividing-line, leaving one daughter-cell with three of a given chromosome and the other with one or, in meiosis, one daughter with two of a particular chromosome and the other with none.
A sex cell with a missing or duplicate chromosome will go on to fuse with another individual's sex cell, which itself will contain a copy of the same chromosome. The resulting zygote will then contain either one or three of that chromosome, and if it survives at all will end up as a very strange organism.
The exception to this is where it is the X chromosome which duplicates, and then fuses with a cell containing a Y. Having two X chromosomes is not harmful - females do it all the time - so the resulting XXY male will be healthy and not in any way obviously malformed; though he will be subfertile and with minor changes e.g. human XXY males tend to be tall and skinny, and may be slow to talk. XXX females are also viable (= able to live).
Note that the most drastic birth-defects are usually due either to gross chromosome abnormalities or to exposure to teratogens (substances which warp an embryo's development without altering its genes, so that the deformation is not heritable) rather than to actual genetic mutations.
Also note that new genetic material can sometimes be introduced not by classic mutation but as a result of viral infection. It can happen that a virus picks up part of the genetic code of its host and incorporates it into its own RNA, then carries that free-floating code to a subsequent host where it may become attached to the new host's genome. If this happens to a sex-cell then the new genetic material becomes heritable: the "blue" (actually a bluish-looking grey) coat-colour in mammals sometimes crops up in new species by this route.
Mutations of Hox genes in insects result in strange aberrations such as legs growing where antennae should be. Vertebrates with Hox gene mutations normally die before birth, but it has been suggested that mutations in Hox genes are involved in major changes such as the loss of the hind-limbs in whales, and that, very early on in animal evolution, Hox genes were responsible for the formation of different phyla. [A phylum is a group of animals with a body-plan very different from that of any other group of animals, such as molluscs, vertebrates etc.: the equivalent groups in plants are called divisions.]
It is thought that Hox mutations may sometimes result in the reappearance of ancestral features which had been switched off - such as a tail occurring in a normally tailless species. The characteristic is suppressed in the first place by a Hox mutation but the code for it is still present. This code is no longer being subjected to selection pressure to keep it accurate, and over the very long term it will become corrupted by random mutations - but it will probably remain readable, and capable of being reactivated by a reverse Hox mutation, for many millions of years. Mice with a Hoxa2 mutation have extra, non-functional jaw-bones - of a reptilian pattern.
Genes can also be sex-limited if they affect a characteristic which is only found in one sex. A female may have a gene affecting the shape of the penis, or a male one regulating pregnancy, but they will have no effect and nothing to affect.
Presumably semi-identical twins could form conjoined twin pairs in the same manner as identical twins. It may also be that the cases mentioned above, where otherwise identical twins differed in some characteristic such as hair colour, were in fact unidentified cases of semi-identical twins.
Even where a set of non-identical twins or triplets are all fully-formed and born alive, they may still have exchanged a few cells early in their development, and so be partial chimaeras.
It is unlikely that a mosaic mutation would result in multiple changes to the finished organism, unless it affects a very major controlling gene. If the organism only has e.g. odd patches of colour or of rexing, or a non-hormonal hermaphroditism (mixture of genders), it's probably a mosaiac: if it has two or more such mixed characteristics it's probably a chimaera.
It could be said that all female mammals are mosaics, because they are made from a mix of two cell-types according to which X-chromasome is switched off.